
![]()
From November 2008 through May 2011, WRC studied the movements and habitat use of Jamaican Boas in their natural home ranges in Windsor.
To do this, project veterinarian, Michael Whittingham DVM (being trained by Dr. Craig Rudolph, USDA-Forest Service, USA, in the photo) surgically implanted 9-gram, temperature-sensitive Holohil radio-transmitters, which enabled us to track the movements of individual animals twice per week (minimum) using a hand-held antenna and receiver.
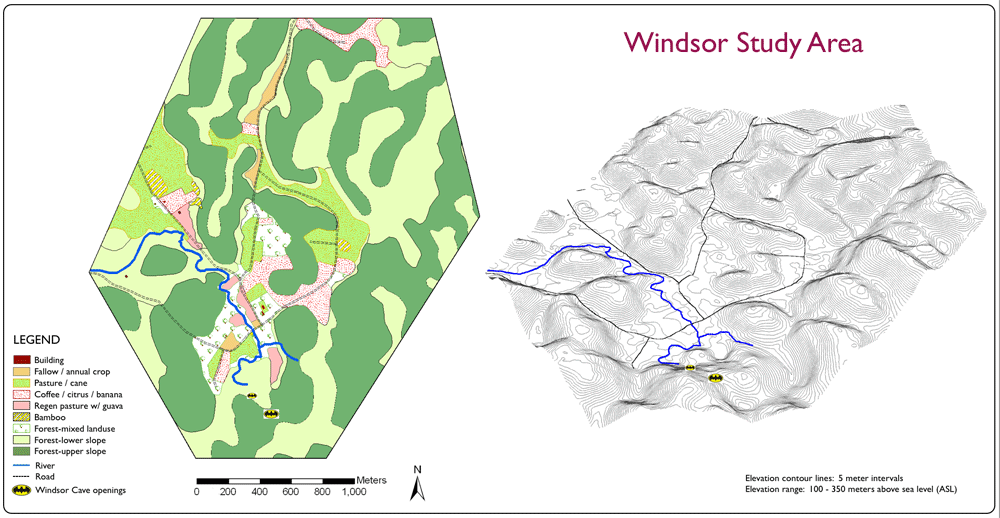
| Macro-habitats available for use by Jamaican Boas. The categories reflect plant species diversity and the physical structure of groundcover, understory density, and canopy coverage. | 3-D Digital Surface Model rotated so viewer angle in 34 0. This enables better visualization
of the cockpit karst morphology.
Click on image to enlarge. |
Although we knew from previous observations that Jamaican Boas move around and actively search for prey (or prey cues) during the night, manpower constraints and safety in the difficult terrain precluded systematic nocturnal assessments. We did, however, opportunistically track a few individuals over multiple nights, in particular a sub-adult male (nicknamed Elvis (aka 00-068B-78D7), who lived in the roof of our guest building. We'll describe nocturnal activities on another webpage.
On this page, WRC shares the results in a series of slides extracted from a Keynote prepared by our director, Dr. Susan Koenig. These include:

| Jamaican Boa moves easily through Rhipsalis cactus. |
| TABLE 1. Introducing the Boas1 Age Class: A=Adult; SA=Sub-adult | ||||||||||||
| Yard Name | ID Code | Weight (g) | SVL (cm) | Age Class | Windsor Cave | Release Date | Transmitter Lifespan (days) | # Fixes | Total Distance Moved (m) | Mean Daily Move (m/fix)2 | Mean Daily Move (m/move)3 | Mean Days Immobile |
| Females | ||||||||||||
| Guango4 | E512 | 2400 | 120 | A | No | 29-Nov-08 | 137 | 35 | 850 | 9.4 | 26.7 | 13.0 |
| Stubby | DAF9 | 2500 | 178 | A | No | 19-Jan-09 | 220 | 55 | 1769 | 7.8 | 11.1 | 3.8 |
| Thingy1 | 9E5A | 1540 | 142 | A | No | 02-Mar-09 | 276 | 65 | 3061 | 15.0 | 20.9 | 8.6 |
| Thingy2 | E7DC | 1040 | 131 | A | No | 02-Mar-09 | 262 | 70 | 2514 | 11.2 | 13.7 | 4.0 |
| Martha | C78B | 2500 | 169 | A | No | 11-Apr-09 | 639 | 186 | 6028 | 10.2 | 17.1 | 10.8 |
| Maddy | 1DD8 | 980 | 115 | A | Yes | 11-Jun-09 | 249 | 70 | 2960 | 14.6 | 20.4 | 4.9 |
| Toop5 | B0F6 | 1790 | 149 | A | No | 31-Oct-09 | 367 | 96 | 5131 | 17.7 | 22.4 | 4.4 |
| Portia | F94A | 2400 | 175 | A | No | 05-Apr-10 | 395 | 124 | 9507 | 29.8 | 38.6 | 8.2 |
| Males | ||||||||||||
| Elvis | 78D7 | 600 | 122 | SA | No | 27-Nov-08 | 799 | 174 | 1997 | 3.3 | 6.2 | 11.4 |
| George | 9475 | 2320 | 173 | A | No | 19-Jan-09 | 395 | 100 | 8015 | 23.6 | 27.1 | 4.8 |
| Zippy | 39FF | 1300 | 139 | A | No | 02-Mar-09 | 201 | 79 | 5367 | 17.3 | 21.8 | 8.1 |
| Marv | 9255 | 1450 | 150 | A | Yes | 05-Sep-09 | 191 | 45 | 1533 | 11.8 | 15.2 | 4.2 |
| Scarv | 95F3 | 810 | 112 | A | Yes | 05-Sep-09 | 247 | 71 | 1796 | 8.9 | 13.1 | 5.6 |
| Bruce | E947 | 2180 | 163 | A | No | 05-Apr-10 | 311 | 89 | 5728 | 21.1 | 31.7 | 7.6 |
|
1An additional 2 females and 1 male were transmittered but they either died (1 female) or
experienced transmitter failure within four months; they are not included here.
| ||||||||||||
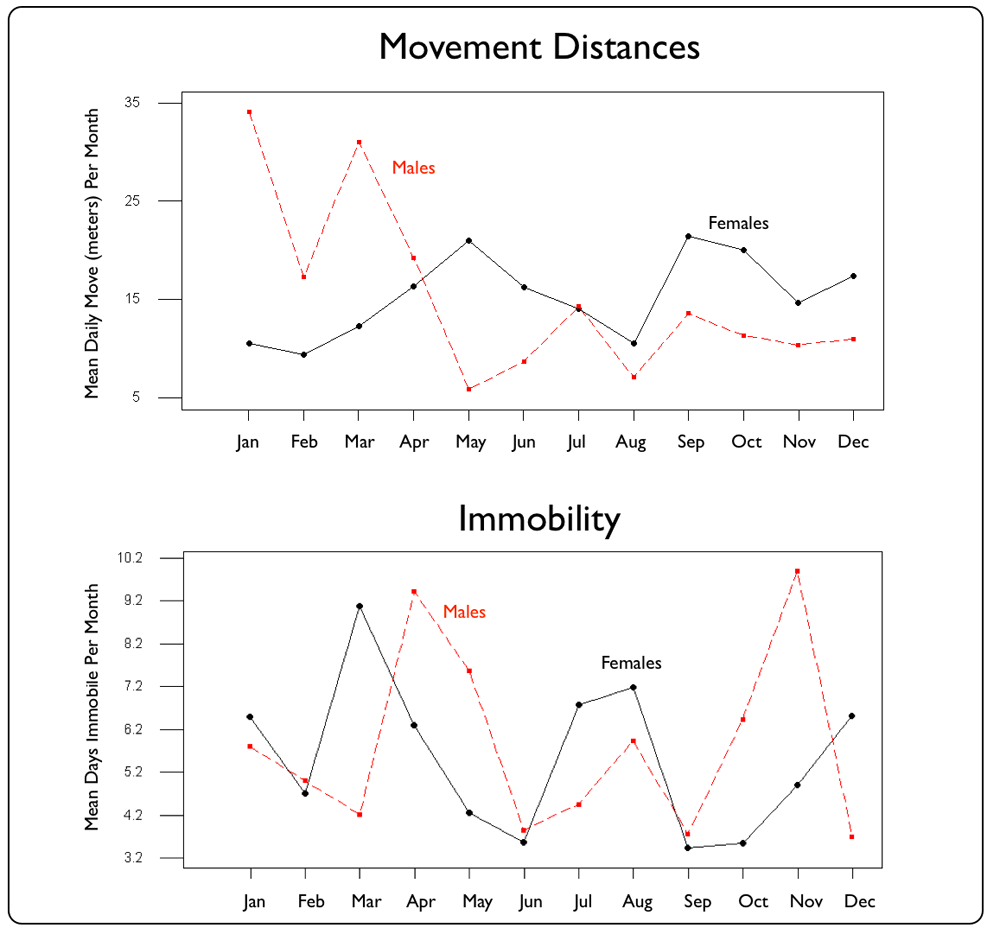
| Jamaican Boas moved an average of 20.4 meters (+ 8.7 SD) daily per move.
Overall, there was no significant (p=0.65) difference in average daily movements between males (19.1 m + 3.9 SE) and females (21.4 m + 3.0 SE) nor in whether a boa was closely associated with Windsor Cave (16.2 m + 2.2 SE) or not (21.6 m + 2.8 SE; p=0.37). Similarly, there was no significant (p=0.82) difference between boas (male & female; cave & non-cave) with regards to the number of consecutive days they remained immobile (females = 7.2 days + 1.2 SE; males = 6.8 days + 1.0 SE). However, these "average" assessments mask both the large distances traveled by males in the first quarter of the year and also the bi-modal peaks in April-May and November, when males were noticeably more sedentary over consecutive days. The April-May peak also corresponds to five observations of pairs of boas either coiled in copulation (23 Apr 2009) or < 5m apart in the same tree (20 Apr 2009;15 & 29 Apr 2010; 26 May 2010). See also Attraction-Avoidance Behaviour. |
 |
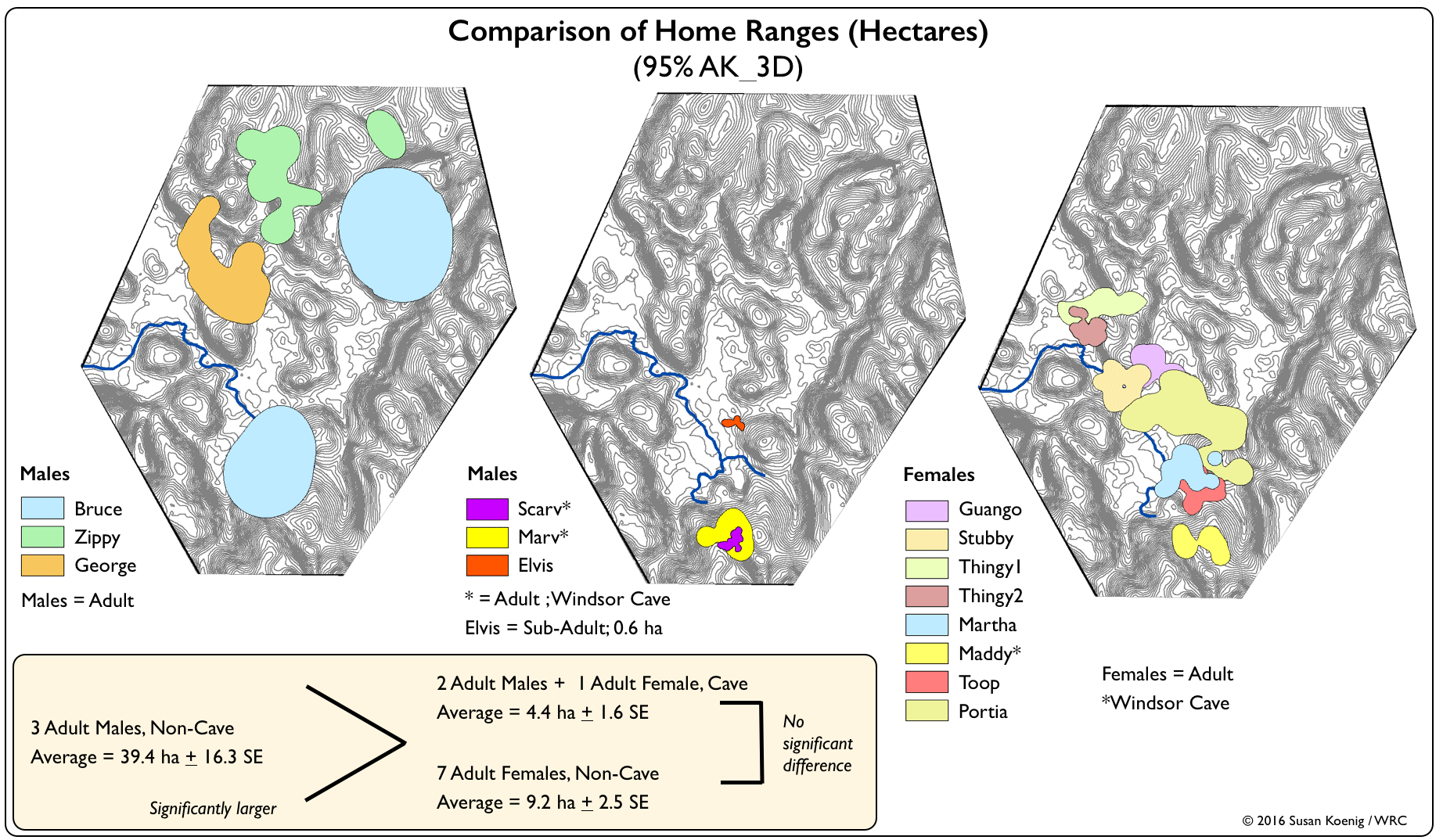
| TABLE 2. Home Range Estimates (Hectares) A home range can be described as the cognitive map (the area an animal knows and maintains in its memory because the area has some value) of where an animal lives (Powell 2000). "Values" include food resources, travel routes, thermoregulation sites, home ranges of members of the opposite sex, etc. Within a home range, if an animal uses some areas with greater intensity, a "Utilization Distribution" can be generated to aid in identifying highly-preferred or biologically-critical resources. For Jamaican Boas, we determined the size, shape and usage intensity of home ranges (95%) and core areas (50%) using the Adaptive Kernel (AK) method. The best reference bandwidth was identified when the 95% isopleth transitioned from encircling discrete patches to becoming continuous (i.e., the "incremental increase" method recommended by Kie et al. 2010). But note, for two males (Zippy & Bruce), their isopleths never merged to a continuous outline because they did big sprints. Where relevant (e.g., attraction-avoidance behaviour), we describe Bruce's "Pre-sprint River Range". We present AK-2D (planar surface area) along with Minimum Convex Polygon (MCP) estimates to facilitate comparison to other studies (e.g., Puente-Rolón and Bird-Picó 2004, Wunderle et al. 2004), but given that planar analyses can underestimate the size of home ranges in topographically-variable landscapes, we also present polygon volume (i.e., 3D) estimates derived from a Triangular Irregular Network (TIN) model. Please email us for full details on methods and analytic techniques.
Click here to see all boa locations on one map or click on any boa's name to view individual maps.
| |||||||||||
| Boa | Age Class | Windsor Cave | # Unique Locations | 50% MCP |
95% MCP |
50% AK_2D | 95% AK_2D | 50% AK_3D | 95% AK_3D | % increase 2D to 3D: 50% AK | % increase 2D to 3D: 95% AK |
| Females | |||||||||||
| Guango | Adult | No | 13 | 2.24 | 4.20 | 1.42 | 9.64 | 1.69 | 10.41 | 16% | 7% |
| Stubby | Adult | No | 35 | 1.14 | 3.60 | 1.08 | 6.55 | 1.27 | 7.40 | 15% | 12% |
| Thingy1 | Adult | No | 40 | 0.64 | 3.60 | 0.70 | 6.53 | 0.78 | 6.98 | 9% | 6% |
| Thingy2 | Adult | No | 48 | 0.70 | 2.16 | 0.57 | 3.12 | 0.62 | 3.34 | 7% | 7% |
| Martha | Adult | No | 82 | 1.27 | 6.04 | 1.13 | 6.79 | 1.13 | 7.28 | 0% | 7% |
| Maddy | Adult | Yes | 36 | 1.12 | 2.51 | 0.81 | 4.78 | 0.88 | 5.16 | 8% | 8% |
| Toop | Adult | No | 56 | 1.21 | 4.89 | 1.01 | 6.37 | 1.08 | 6.93 | 6% | 8% |
| Portia | Adult | No | 83 | 6.63 | 19.56 | 4.09 | 22.04 | 4.26 | 23.53 | 4% | 6% |
| Males | |||||||||||
| Elvis | Subadult | No | 42 | 0.07 | 0.65 | 0.02 | 0.59 | 0.02 | 0.61 | 0% | 4% |
| George | Adult | No | 67 | 0.94 | 16.28 | 1.88 | 20.53 | 2.37 | 23.04 | 21% | 11% |
| Zippy | Adult | No | 47 | 0.50 | 21.63 | 0.77 | 20.88 | 0.89 | 23.00 | 13% | 9% |
| Marv | Adult | Yes | 30 | 0.34 | 2.25 | 0.63 | 5.92 | 0.69 | 6.71 | 8% | 12% |
| Scarv | Adult | Yes | 31 | 0.20 | 0.75 | 0.16 | 1.10 | 0.18 | 1.22 | 13% | 12% |
| Bruce | Adult | No | 38 | 0.93 | 70.11 | 2.81 | 64.95 | 2.85 | 71.96 | 1% | 10% |
 |
|
Why are cave-associated home ranges so small?
Windsor Great Cave, which is owned by WWF-UK and managed by WRC, supports one of the largest and most species-diverse bat roosts on Jamaica. There are more than 100,000 bats, of 11 different species, which emerge at dusk for their nighttime feeding activity throughout the wider landscape. Taking advantage of Strangler Fig (Ficus spp) roots growing along the rock face, a boa can hold on by its lower body and dangle vertically in the huge stream of emerging bats. Immature boas strike at everything they detect . . . and miss almost everything. With experience, they learn to hold their strike until a bat actually makes physical contact with them . . . and then, occasional success. Because the bats emerge every night (unless it's raining) and their flight activity is timed to solar sunset cues, they are an abundant, concentrated, and reliable prey source year-round. It is well worth a boa's effort to stay in close proximity to a bat-occupied cave. Indeed, the highest densities of Chilabothrus boas in the Greater Antilles are recorded at bat caves (Rodríguez-Cabrera et al. 2015 and references therein).
|
 |
|
Were boas attracted or
did they avoid other?
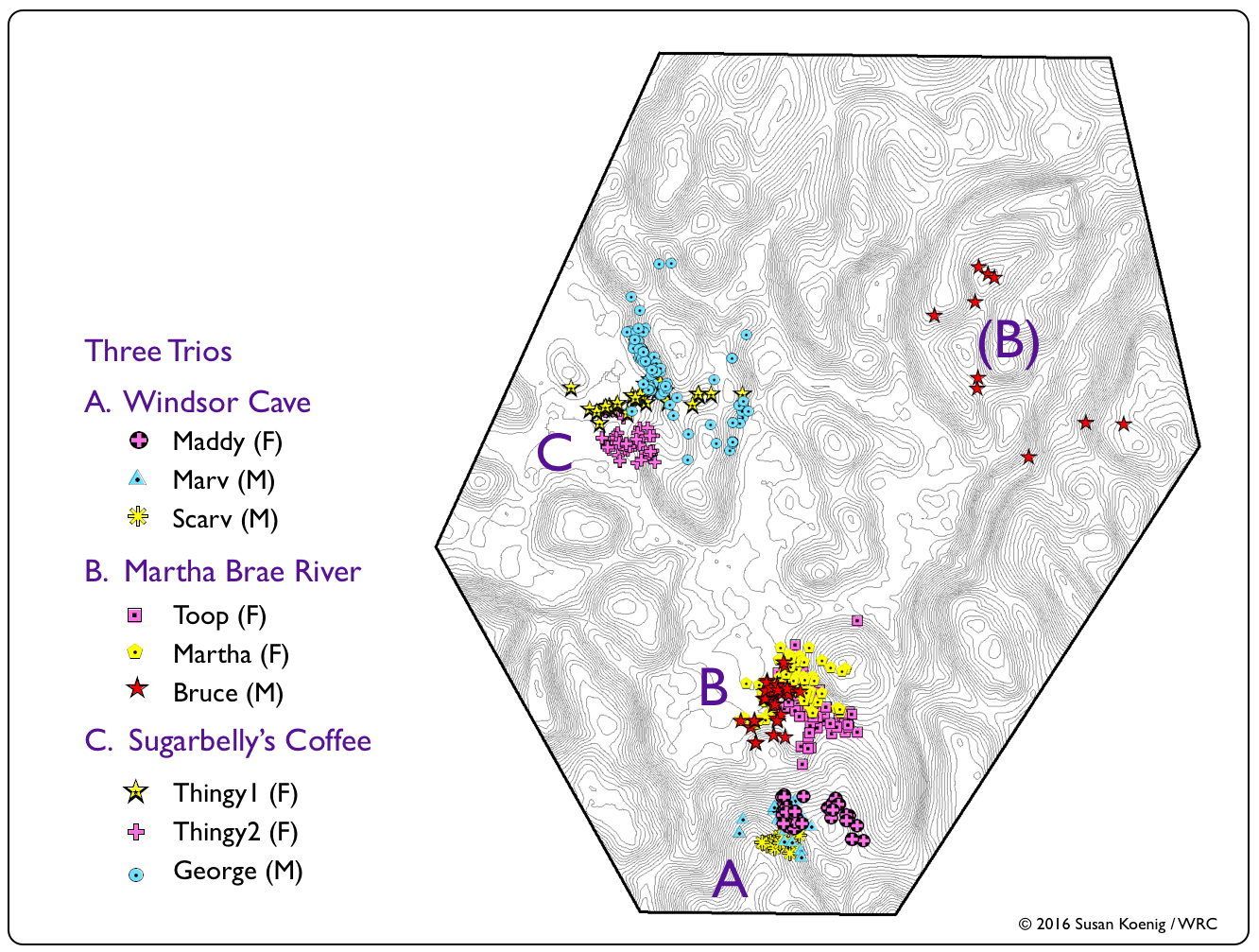
With the spatial co-occurrence of so many snakes, we wondered whether they would continually run (slither...) into each other (i.e., demonstrate some degree of tolerance or attraction) or if they maintained a minimum distance (i.e., avoidance behaviour) in their spatially-overlapping ranges. To explore the likelihood of boas encountering each other, we first examined the percentage of overlap in their 95% AK home ranges on a monthly basis (static relationships) and then examined the distances they maintained between daily fixes (dynamic interactions). |
Spatial vs. Temporal Overlap Zones (OZ)
|
Windsor Cave Trio
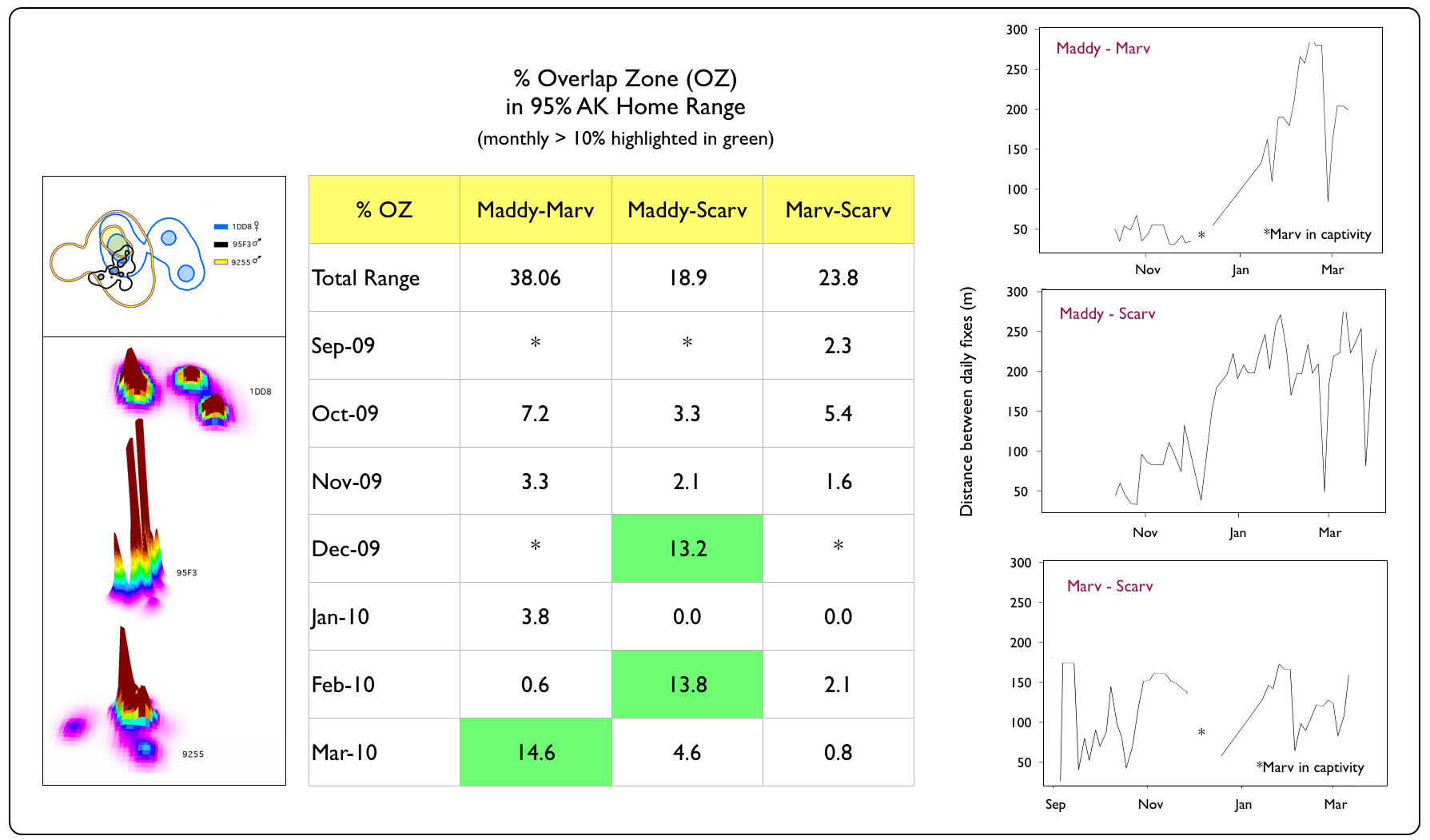
Dynamic interaction distance graph generated using wildlifeDI package (Long 2014) for R software v3.2.3 (The R Foundation for Statistical Computing 2013). |
Martha Brae River Trio
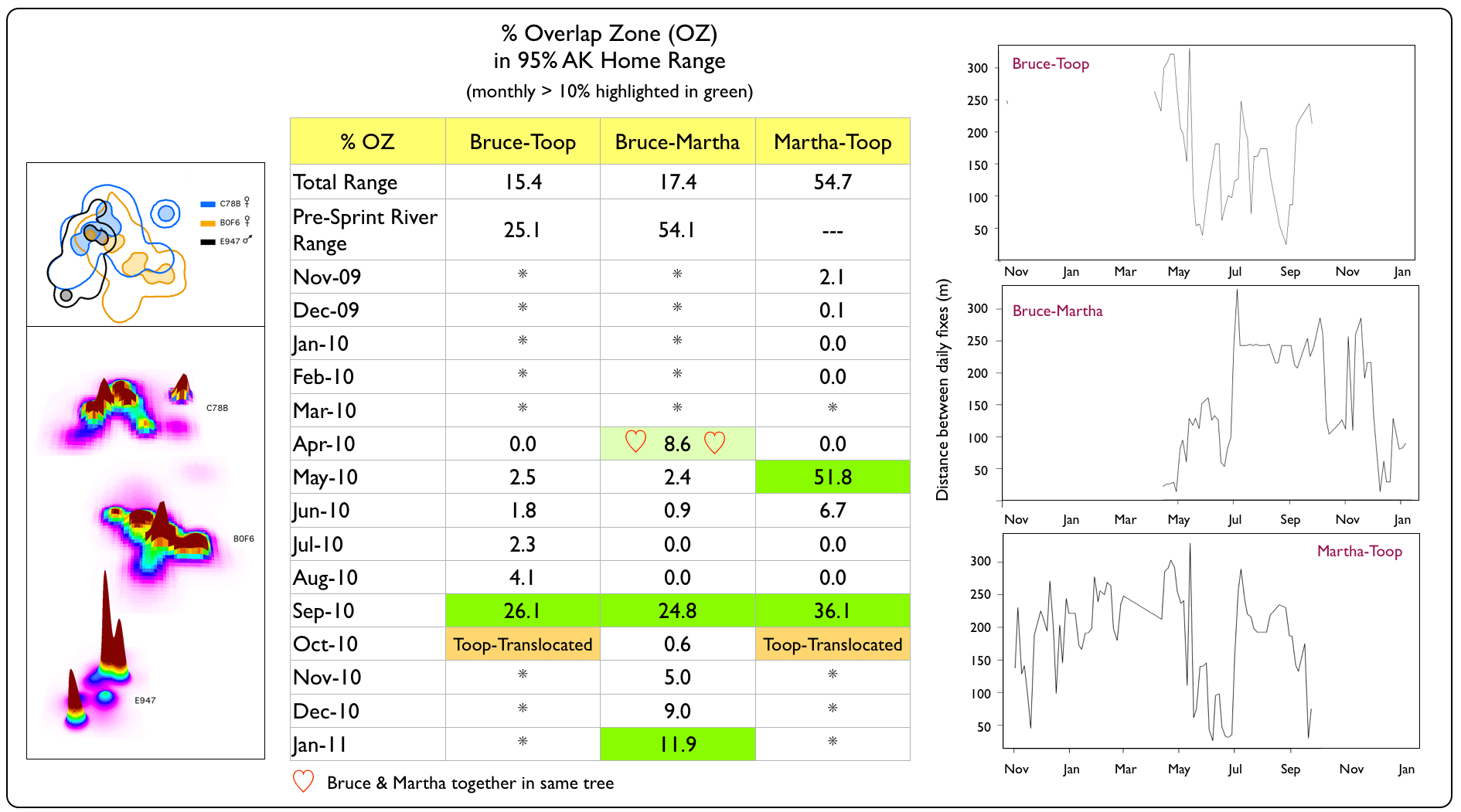
Dynamic interaction distance graph generated using wildlifeDI package (Long 2014) for R software v3.2.3 (The R Foundation for Statistical Computing 2013). |
Sugarbelly's Coffee Trio
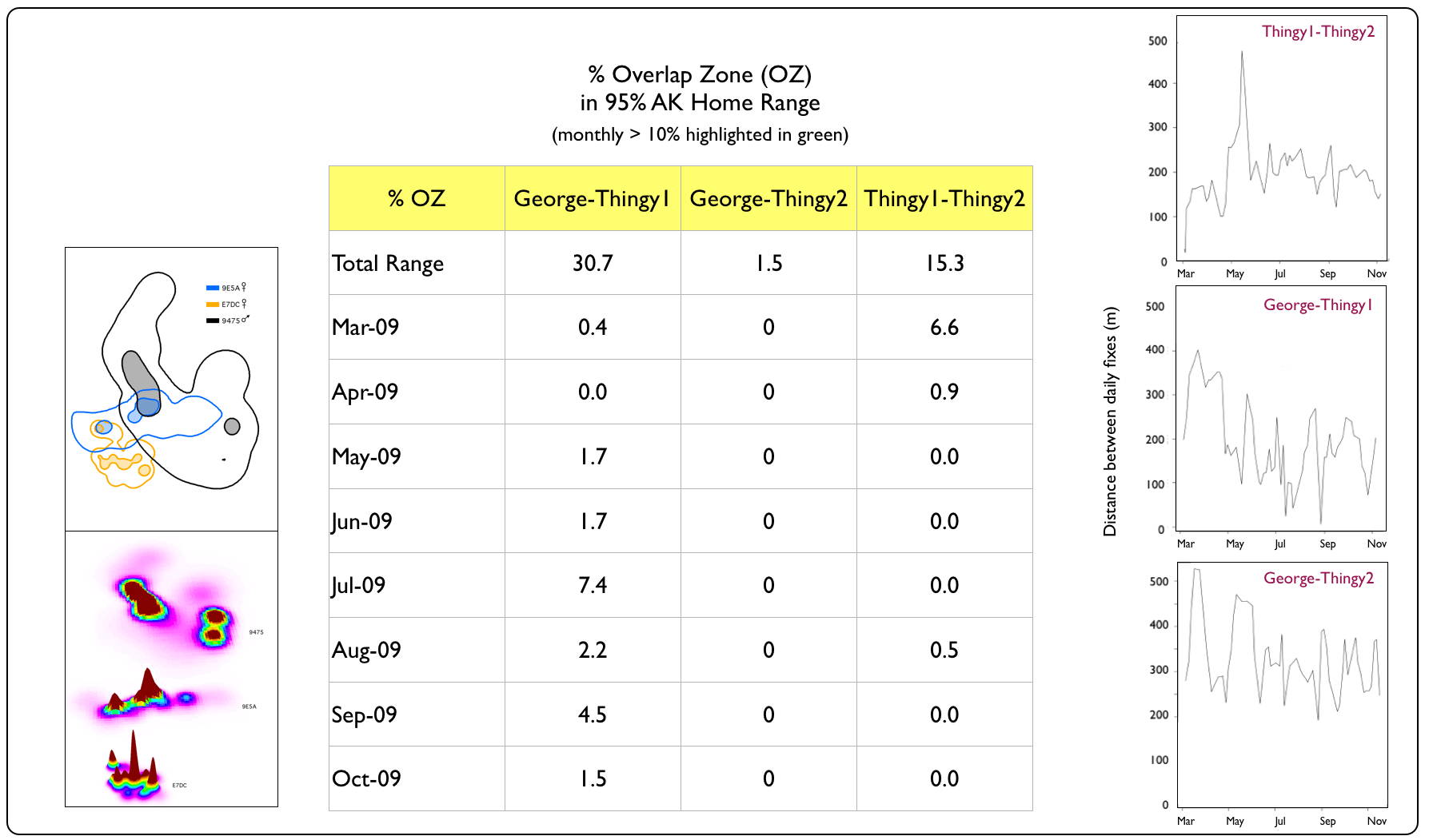
Dynamic interaction distance graph generated using wildlifeDI package (Long 2014) for R software v3.2.3 (The R Foundation for Statistical Computing 2013). |
|
Were boas attracted or
did they avoid other?
Despite having spatially-overlapping territories (esp. the Windsor Cave and Martha Brae River trios), Jamaican Boas appeared to avoid each other in their home ranges for most of the year. Notable exceptions were in April 2010, when we captured Bruce (non-transmittered at the time) and Martha (transmittered) in the same tree; Marv and Scarv (both non-transmittered) in September 2009, resting on the same horizonal snag at the Windsor Cave opening; and Thingy1 and Thingy2 (both non-transmittered) in March 2009, resting in coffee trees. Although the Windsor Cave boas maintained the closest distances of all trios, they also demonstrated the ability to avoid each other while resting during the daytime. Such behaviour may enable boas to reduce the chances of being detected by potential prey items or by the parents of potential prey items (e.g., adult birds protecting their nesting territory & nestlings, such as we see with these Jamaican Crows). |
Attraction / Avoidance Behaviour
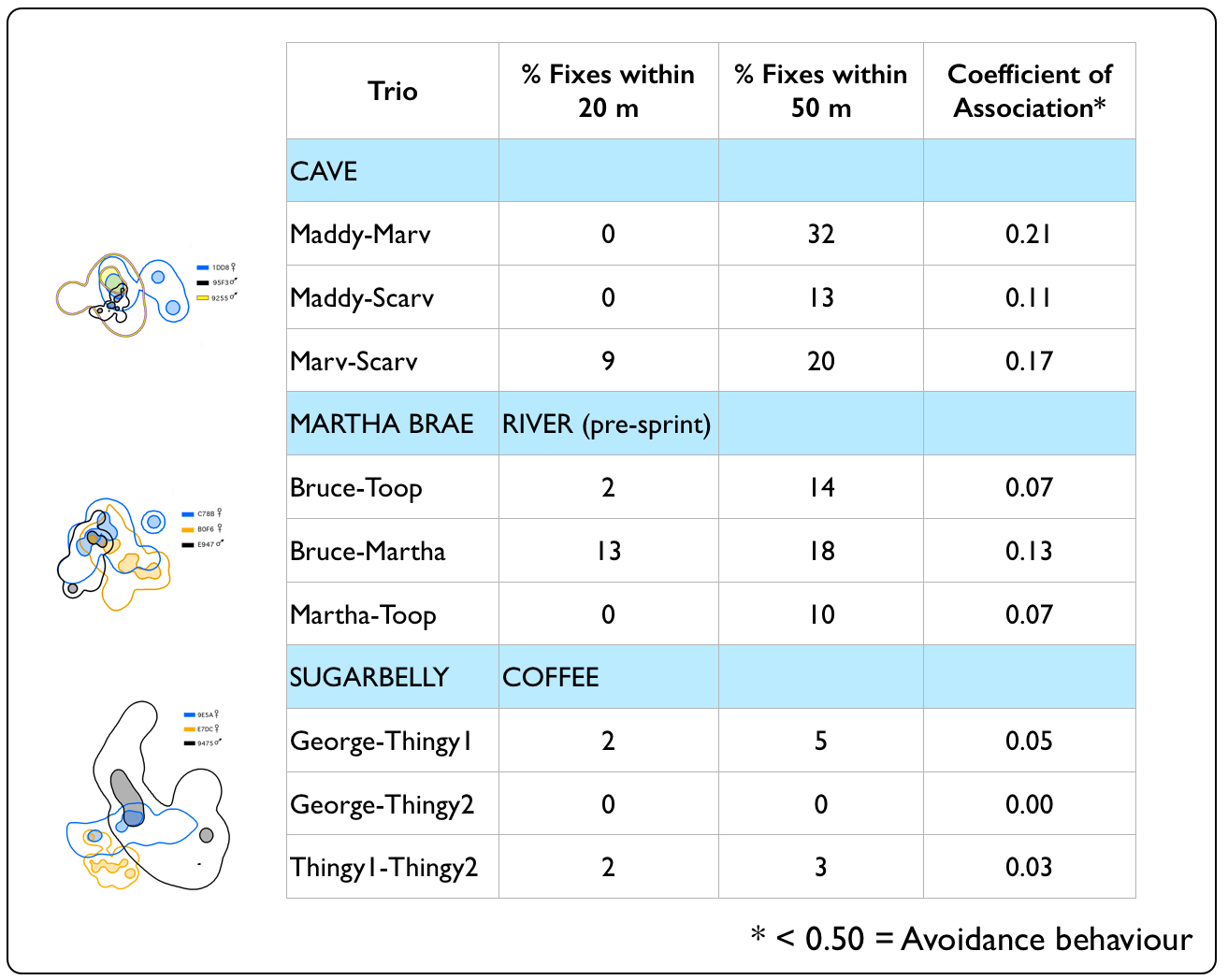
Distance percentages and Coefficient of Association generated using wildlifeDI package (Long 2014) for R software v3.2.3(The R Foundation for Statistical Computing 2013). |
|
We didn't really need fancy statistical software to tell us that Jamaican boas loved to use cliffs
more than one would predict based on availability. Ugh, "George" was a nightmare to track.
Although boas used relatively flat areas (i.e., slope < 200) as one would expect from availability, there is a subtlety to note: in cockpit karst, both bottomlands and conical hilltops are gently sloped (it's always a relief to get to the top!). But we see that they preferred to stay at lower elevations (except for the boas at the top opening of Windsor Cave [elev. 220m]) and they used "Forest-Mixed Landuse" (i.e, immature forest with old cultivated fruit trees) significantly more than one would have expected. This was the most structurally-complex of the landuse categories. Forested bottomlands are important! We also note that boas signifcantly avoided the most open of agriculture areas, esp. yam farms and sugar cane / pasture. |
Macro-Habitat Features Type I: Population-Level Preferences 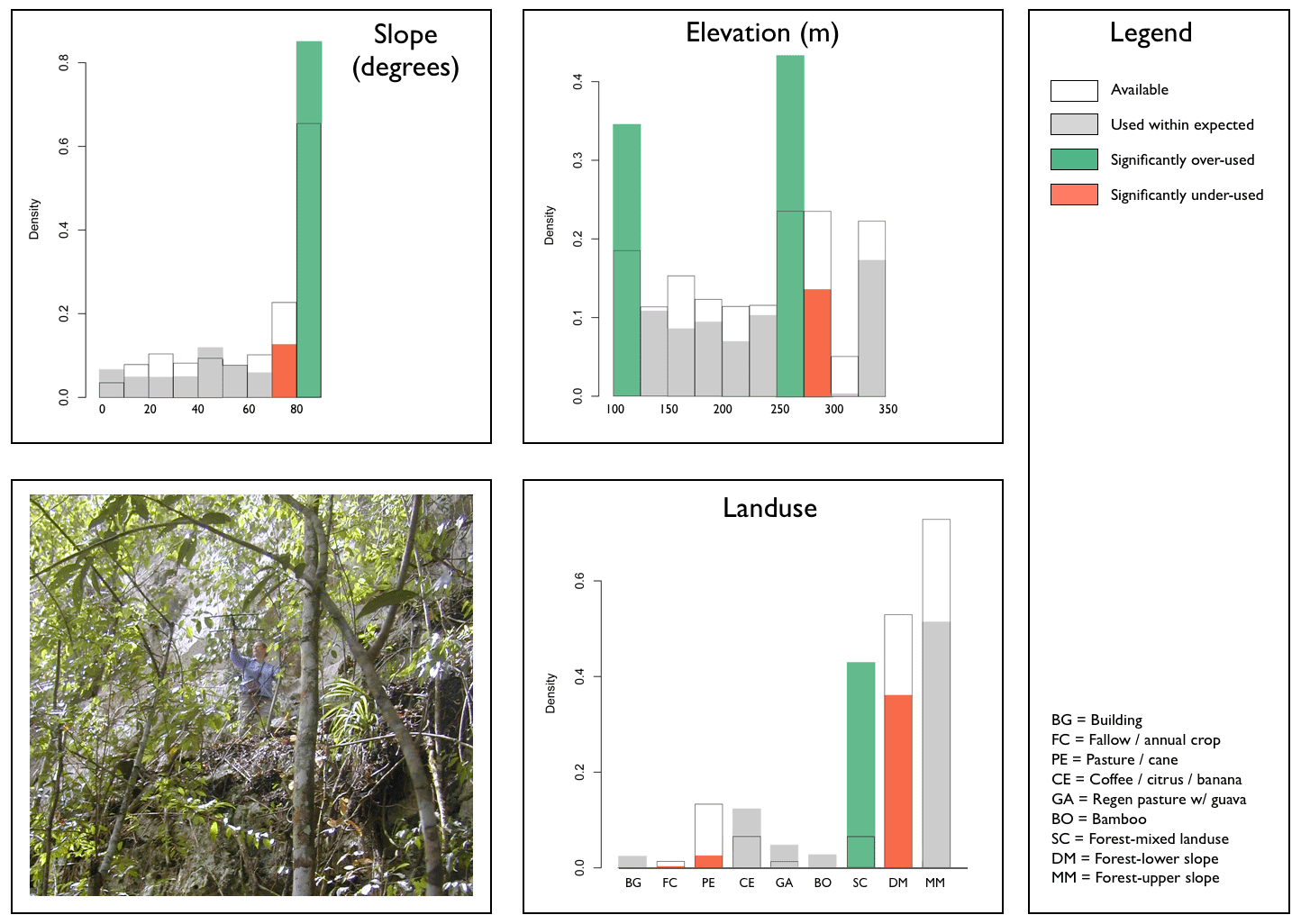 |
| Although the boas' choice of aspect was over-used at 30-600, and their aspect choices, statistically, were not uniformly distributed, there's not an immediately obvious pattern to make us say "wow, it looks like they were trying to position themselves to thermoregulate or to keep themselves oriented for solar or celestial cues...or whatever." Interpretation of aspect isn't straightforward in cockpit karst. For example, the southern exposure of one hillside might actually be in shadow because of a taller adjacent hill. |
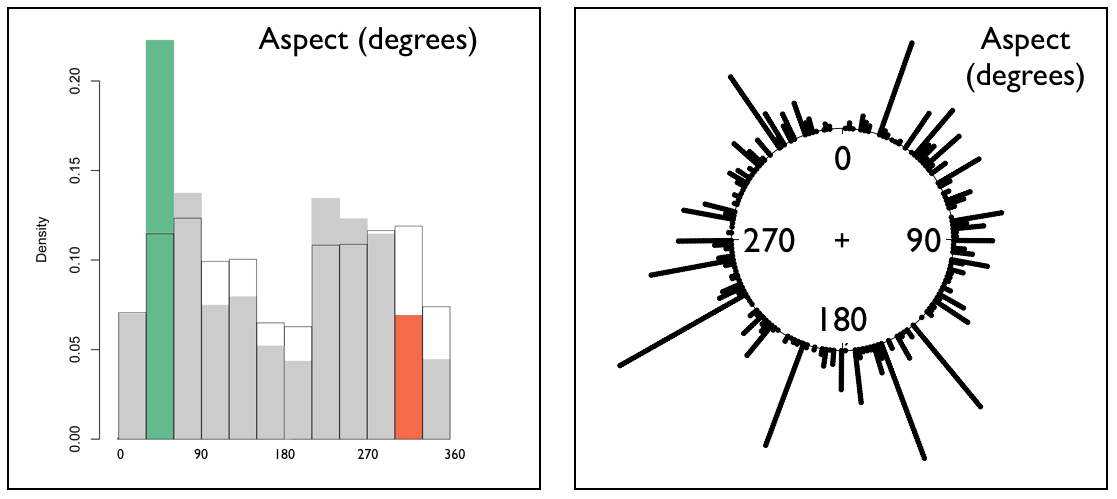
Histograms and statistical results computed using adehabitatHS package (Calenge 2011) for R software v3.2.3 (The R Foundation for Statistical Computing 2013). Colour denoting significant results added by Koenig. |
| Because of the landuse patterns, which are strongly associated with topography, all boas theoretically had short-distance access to multiple landuse categories. For example, the boas at Windsor Cave were within 200-250m of bottomland habitat (a pasture regenerating with guava trees). However, these three boas chose to remain near the top cave opening. Consequently, and unlike all of the other boas, they associated almost entirely with the closed-canopy, upper-slope forest type. |
Macro-Habitat Features Type II: Individual Preferences 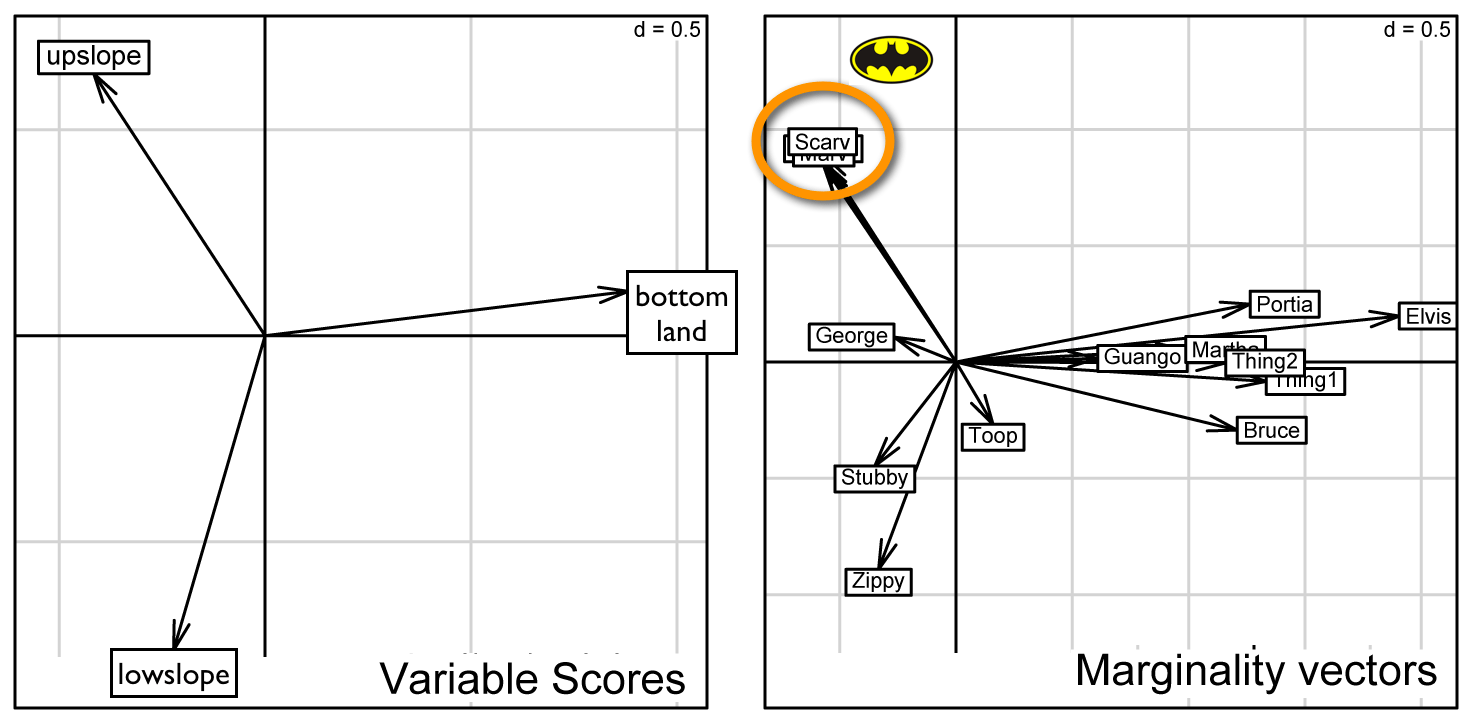 |
|
For those mature readers:
Elvis was in the building!
But as you can see, Elvis' strong love of spending his days in the roof of "his" outbuilding overwhelmed the dataset and made it difficult to visualize the preferences of the other adult boas. So, we deleted sub-adult Elvis' data . . . |
Individual Preferences (incl. Elvis) 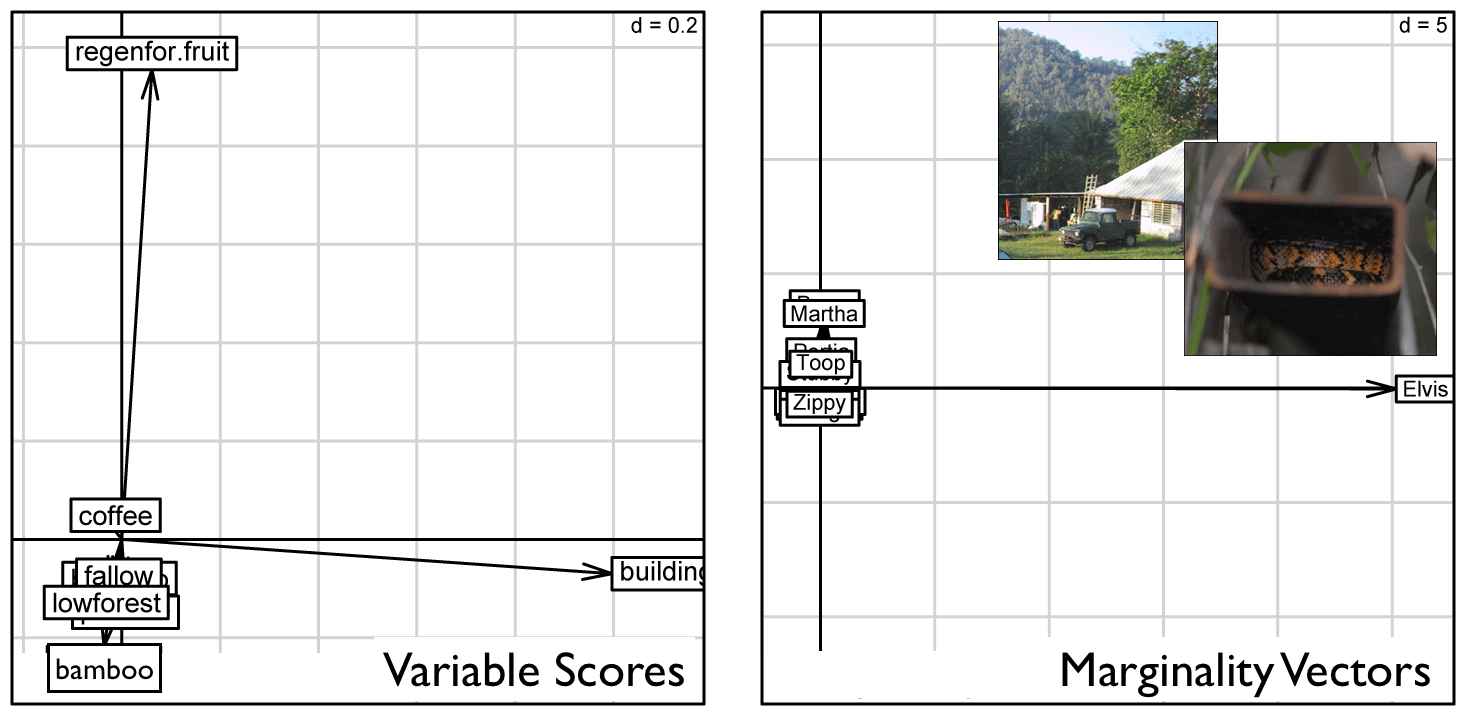
|
| . . . and now we can see the individual adult boas that showed strong preferences for the structurally-complex bottomland landuse of "regenerating forest with old cultivated fruit trees" and the slightly less complex bottomland agriculture habitat of shade coffee, citrus, and banana but not fallow fields and pastures. |
Individual Preferences (without Elvis) 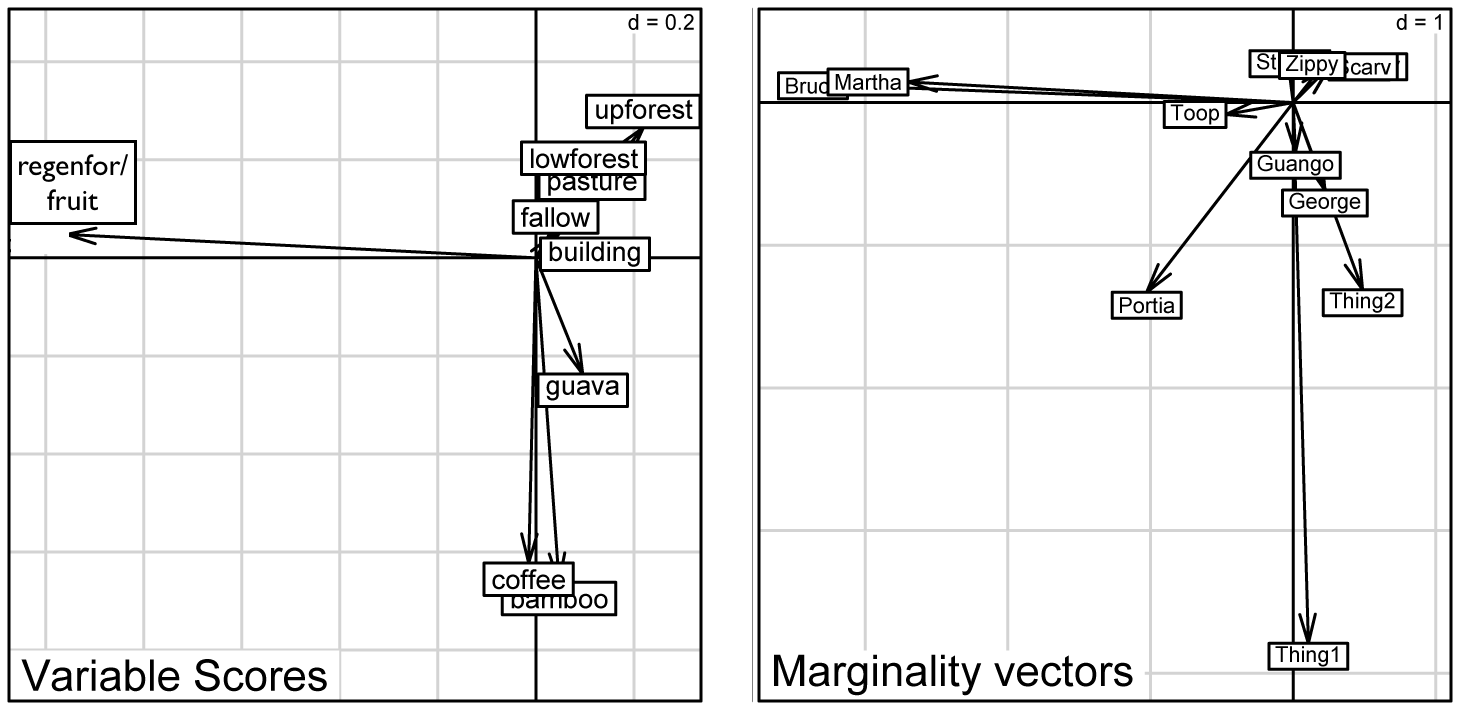
|
| Graphical results of a Canonical Outlying Mean Index (CANOMI) analysis generated using adehabitatHS package (Calenge 2011) for R software v3.2.3 (The R Foundation for Statistical Computing 2013). |
|
Although adult boas used a variety of microhabitats throughout the landscape, live broadleaf trees were the
predominantly used feature (40.1% + 49.0 SD of fixes), followed by ground or below-ground (e.g., rock rubble) sites (19.1% + 36.9 SD) and
vine-choked trees (10.1% + 30.1 SD).
For 187 (176 arboreal and 11 terrestrial) of the 1241 (15%) analyzed locations, boas were either seen amongst large tank bromeliads (Hohenbergia spp.) or the transmitter signal was highly directional to a cluster of bromeliads. These epiphytic plants collect water in the base of their leaves, which in turn attracts many animals to drink or feed on small aquatic invertebrates: bromeliads are to Cockpit Country what watering holes are to predators in the Serengeti. |
Micro-Habitat Preferences
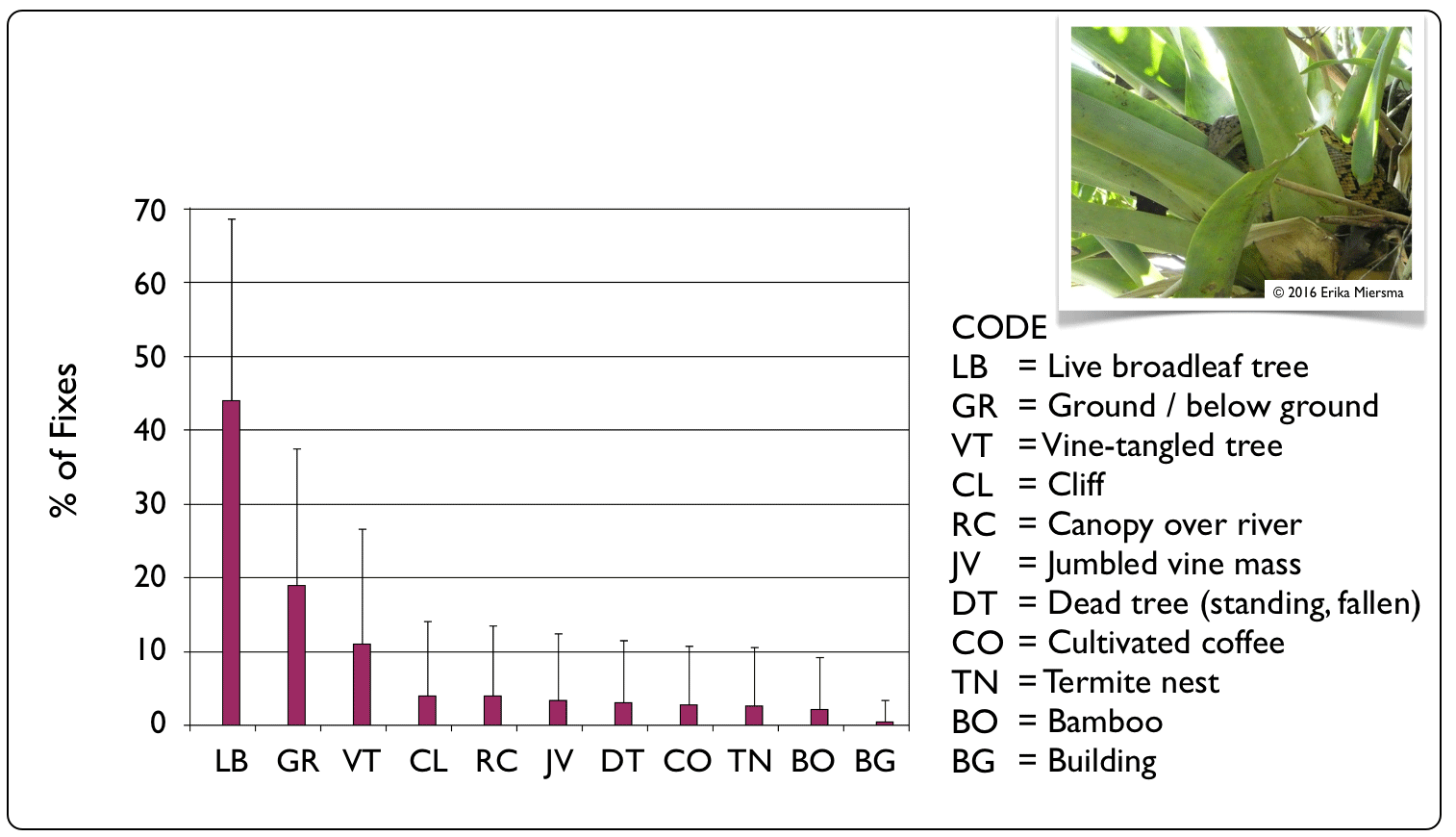
|
| Arboreal vs. Terrestrial Excluding data from inaccessible cliffs (where we couldn't confirm whether the boa was tucked away in a ledge or in a tree growing in the rock face; 5% of the total number of fixes), boas displayed a significant (p < 0.001) preference for arboreal daytime resting locations: 73% + 15 SD of fixes were arboreal. The trend for males (76% + 6 SE ) to be located more often arboreally than females (71% + 6 SE ) is on the edge of being statistically significant (p = 0.054). |
|
Micro-site fidelity With the exception of "Guango", who was tracked for the shortest period of time (~ 4 months), all boas revisited previous location fixes. Females returned to 10% of locations one time, 2% of locations two times, 1% of locations three or four times, and <0.5% of locations 5, 6, or 7 times. Revisitation of sites occurred most often in June and August-November. Adult males showed similar rates of revisitation, with 15% of locations revisited once, 2% revisited twice, 1% revisited thrice, 2% revisited four times, 1% revisited 5 or 6 times, and < 0.5% revisited 7 times. In contrast to females, they least often revisited sites in September-October and most often revisited sites in December. Sub-adult male "Elvis" has a different story. The outbuilding was the centre of his universe: 60% of his daytime resting fixes were in the roof of the building. Within the roof, he particularly liked to return to the SW corner of Room #1, which had good connectivity to an adjacent ackee tree (a tree in which he rested on 4 different occasions during the day and visited on 13 different nights, including one night to eat the nestlings of our "yardie" Jamaican Crow pair 😢 ) and the east-facing part of the roof over Room #3, over the door... so visitors should still keep an ear open for him! To keep himself oriented on the eastern edge of his home range, he liked a huge Strangler Fig tree, which he visited 6 times. We have much more to say about Elvis, so stay tuned. |
|
| Visual Detectability Jamaican Boas are incredibly cryptic: even when we were within a few meters of a transmittered animal, we only observed them in 20.2% (+ 14.0 SD) of the fixes. Although there was a trend for females to be more visible (25.1% + 0.05 SE) than males (14.7% + 0.05 SE), this was not significant (p = 0.14). Interestingly, although there was no significant (p = 0.19) interaction by month for females, males were significantly (p = 0.05) more likely to be seen in the month of August.
Rare...or Difficult to Detect? Our limited success was not because Jamaican Boas are rare in Windsor; on the contrary, we'd have to say that they are as common as dirt here! But if someone said "do a specific search and capture survey" to find a Jamaican Boa or to estimate the population size and density, our default setting would be "we're going to seriously underestimate the number present if you ask us to do a Rapid Ecological Assessment." |
 Can you find the boa? Click on the picture or here for a short video. |
| TABLE 3. Characteristics of Trees Used by Jamaican Boas We present those characteristics of boa trees which were significantly different (p value < 0.05) from trees sampled randomly at a distance of 20 meters from the boa's location. Vines clearly make it easy for boas to move around their microhabitats. The percent-connectivity (mean + SE) of a boa's tree with its neighbouring trees was almost significantly different from random trees (boa tree connectivity = 47.8% + 3.4 SE VS. random tree connectivity = 39.7% + 2.6 SE; p=0.055). Click here for the complete list of habitat variables assessed. |
|||
| Tree Variable | Boa Plot Mean + SE |
Random Plot Mean + SE |
P-value | Tree height (m) | 19.3 + 1.3 | 14.8 + 0.7 | 0.005 | Tree DBH (cm) | 43.8 + 3.9 | 21.1 + 1.8 | 0.000 | Cavity in bole or branch (% of trees with) | 26.7 + 2.1 | 8.4 + 2.0 | 0.000 | Vine species diversity: number of different species on bole | 2.0 + 0.1 | 1.3 + 0.1 | 0.000 | Height of vines attached on bole (m) | 14.5 + 0.9 | 9.4 + 0.5 | 0.000 | Percentage of tree height of vines attached on bole | 76.6 + 2.1 | 64.3 + 4.3 | 0.016 | Density score of vines attached on bole (range 0 - 3) | 1.8 + 0.1 | 1.2 + 0.1 | 0.001 | Vine species diversity: number of unattached vine species | 2.0 + 0.1 | 1.3 + 0.1 | 0.000 | Height of unattached vines (m) | 12.5 + 0.6 | 9.1 + 0.7 | 0.001 | Density score of unattached vines (range 0 - 3) | 1.6 + 0.1 | 1.0 + 0.1 | 0.001 | Medium & large1 bromeliads (number) | 5.2 + 0.6 | 1.0 + 0.2 | 0.000 | Ferns (number) | 2.6 + 0.3 | 0.6 + 0.1 | 0.000 | Termite nests (number) | 0.1 + 0.02 | 0.0 + 0.00 | 0.000 |
| 1Medium and large bromeliads provide both "climbing structure" and aquatic reservoirs.
Small bromeliads, which can provide a "surface grip" but no appreciable reservoir, did not differ between boa and random trees.
PHOTO: Termite (Nasutitermes sp.) nest being used by Jamaican Parakeets (Eupsittula nana) for nesting in Windsor, May 2009. Jamaican Boas are confirmed predators of parakeets (Koenig et al. 2015). In addition to providing feeding opportunities, termite nests also are used by adult female (probably gravid) boas, possibly for thermoregulation (this study). |
 |
||
Acknowledgements:
| Funding | Equipment | 
Southern Research Station Nacogdoches, TX |
 |
 Wildlife Without Borders Latin America and the Caribbean |
 |
WRC extends an enormous thank you to D. Craig Rudolph (USDA-Forest Service [USDA-FS]). Craig co-conceived the project with WRC's Susan Koenig, trained the project's veterinarian, Michael Whittingham, DVM, provided on-going advice, secured funding from USDA-FS which kept the project running from start-to-finish, and kept the project equipped with functional radio receivers. This latter was no small undertaking in the high humidity and afternoon thunderstorms of Windsor. Thanks to Craig and USDA-FS, we didn't miss any field days because of dead equipment.
Thanks also to other USDA-FS personnel who helped launch the project: Josh Pierce, Rick Schaefer & Howard Williamson. Dino Ferri (Jacksonville Zoo and Gardens, Florida; now at the Central Florida Zoological & Botanical Gardens) provided equipment and gave us some of his time for field tracking. Paul Hime (now at the University of Kentucky) also helped with the launching of the project.
We also thank Michel C. Milinkovitch for donating Trovan ID-100B RFID transponders and the associated Trovan Reader: this ensured that we could identify uniquely all boas we encountered.
Erika Miersma (US Peace Corps volunteer) was invaluable for keeping the project running in the first year. We also thank field assistants Elodie Gago, Marvin Grossett, "Hoggy", Zain Mallett and Todd Stimpson Alleger.
Thanks also to Mathieu Basille for the coding fix for the "count.point" function in adehabitatHS - this enabled us to run the Type II habitat preference analyses.
Finally, we thank all of the Windsor farmers who reported boas to us. They appreciated there was value in not killing a boa on-sight.
Citing information on this webpage:
Koenig, Susan E. Jamaican Boa home range, attraction-avoidance behaviour, and habitat preferences in Cockpit Country, Jamaica. Windsor Reearch Centre, 2016-17. Retrieved from www.cockpitcountry.com/JamaicanBoaTelemetry.html, DATE DOWNLOADED.
Please email us with any questions you have.
Literature & Software:
Calenge, C. 2011. Exploratory Analysis of the Habitat Selection by the Wildlife in R:
the
adehabitatHS Package. On-line manual.
For software package: adeHabitatHS
ESRI ArcMap v9.3 Geographic Information System (GIS) software. Environmental Systems Research Institute, Redlands, California, USA. Esri.com.
Kie, J. G., J. Matthiopoulos, J. Fieberg, R. A. Powell, F. Cagnacci, M. S. Mitchell, J-M. Gaillard, and P. R. Moorcroft. 2010. The home-range concept: are traditional estimators still relevant with modern telemetry technology? Philosophical Transactions of the Royal Society B 365: 2221–2231.
Koenig, S., C. Levy, and V. Turland. 2015. Jamaica's endemic parakeet Eupsittula [formerly Aratinga] nana (Vigors, 1830): Natural history and conservation status. The Broadsheet 100: 6-18.
Long, J.A. 2014. wildlifeDI: Analysis of Dynamic Interaction Patterns from Wildlife Tracking Data Version 0.2. On-line manual.
For software package: wildlifeDI
Miersma, E. E. 2010. Movements, activity range, habitat use, and conservation of the Jamaican (Yellow) Boa, Epicrates subflavus. Masters thesis, University of Montana, Missoula. Pp. 67. [NOTE: Miersma's analyses were of a subset (November 2008 - November 2009) of the complete baseline dataset summarized on this webpage.]
Powell R. A. 2000 Animal home ranges and territories and home range estimators. In Research technologies in animal ecology—controversies and consequences (eds Boitani L., Fuller T. K.), pp. 65–110. New York, NY: Columbia University Press.
Puente-Rolón, A.R. and F.J. Bird-Picó. 2004. Foraging behavior, home range, movements and activity patterns of Epicrates inornatus (Boidae) at Mata de Plátano Reserve in Arecibo, Puerto Rico. Caribbean Journal of Science 40: 343-352.
Rodgers, A.R. and J. G. Kie. 2011. HRT: Home Range Tools for ArcGIS User's Manual, Draft August 10, 2011. Centre for Northern Forest Ecosystem Research, Ontario Ministry of Natural Resources. On-line manual
Rodríguez-Cabrera, T., J. Torres, and R. Marreno. 2015. At the lower size limit of snakes preying on bats in the West Indies: the Cuban Boa Chilabothrus angulifer (Boidae). IRCB Reptiles & Amphibians 22: 8-15.
The R Project for Statistical Computing software. R version 3.2.3 (2015-12-10). The R Foundation, www.r-project.org
Venables, W.N., D.M. Smith and the R Core Team. 2015. An Introduction to R, version 3.2.3. (2015-12-10). On-line manual.
Wunderle, J.M., Jr., J.E. Mercado, B. Parresol, and E. Terranova. 2004. Spatial ecology of Puerto Rican Boas (Epicrates inornatus) in a hurricane impacted forest. Biotropica 36: 555-571.