

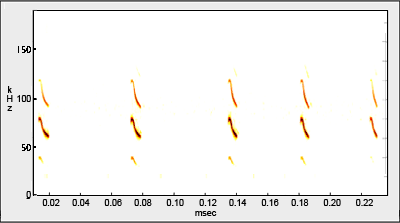

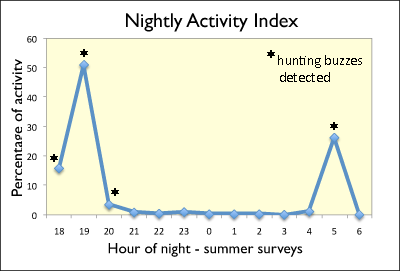
Additional:
Pteronotus quadridens is considered to be the least common Mormoopidae on the island and is currently
known to reside in only 10 caves (NEPA 2011). It has a preference for extensive deep wet cave systems and large high-domed chambers, well away from the cave entrance.
In Cuba and Puerto Rico, pregnant females are found from February through June, with a peak in May when parturition begins. While some females may still be lactating into September, July is when most females are suckling pups (Gannon et al. 2005; Silva Taboada 1979). In Jamaica, Goodwin (1970) reported that none of the 15 adult females captured in mid- to late July showed any evidence of reproductive activity, so breeding may commence earlier in Jamaica . . . but we urge caution drawing conclusions from the very small sample sizes.
In Cuba, diet is dominated by beetles (Coleoptera) and flies (Diptera). Moths (Lepidoptera), katydids, crickets & grasshoppers (Orthoptera) and true bugs (Hemiptera; incl. previously recognized Homoptera) are consumed less frequently (Silva Taboada 1979). Spiders (Araneae) are consumed on rare occasions (Mancina et al. 2012). In Puerto Rico, diet was found to be dominated by Coleoptera, Lepidoptera, and Hymenoptera (bees, wasps, ants); Hemiptera and Diptera were consumed less frequently, and Orthoptera and Araneae only rarely (Rolfe 2011).
Pollen grains in fecal samples and stomach contents of P. quadridens have been reported both in Cuba (Silva Taboada 1979, Mancina et al. 2012) and Puerto Rico (Rodríguez-Durán and Lewis 1987). Although pollen may have been carried by insect prey, it also appears that P. quadridens is capable of capturing insects while in hover-flight at flowers (Mancina et al. 2012).
Acknowledgements:
Data presented on the Windsor Great Cave colony were collected through the support of:
| |
|
|
Literature Reviewed
Gannon, M.R., A. Kurta, A. Rodriguez-Duran, and M.R. Willig. 2005. Bats of Puerto Rico. University of the West Indies Press, Mona, Jamaica. pp. 239.
Genoways, H.H., R.J. Baker, J.W. Bickham, C. J. Phillips. 2005. Bats of Jamaica. Museum Texas Tech Univ. Texas, USA. pp 155.
Goodwin, R.E. 1970. The ecology of Jamaican bats. Journal of Mammalogy 51:571-579.
Mancina, C.A., L. Garcia-Rivera, and B.W. Miller. 2012. Wing morphology, echolocation, and resource partitioning in syntopic Cuban mormoopid bats. J. Mammology 95: 1308-1317.
National Environment and Planning Agency. 2011. Bat Management Plan for Jamaica 2012 – 2017. Ecosystems Management Branch, NEPA, Government of Jamaica.
Rodríguez-Durán, A. and A.R. Lewis. 1987. Patterns of population size, diet, and activity time for a multispecies assemblage of bats at a cave in Puerto Rico. Caribbean Journal of Science 23:352-60.
Rolfe, A. K. 2011. Diet of Three Mormoopid Bats (Mormoops blainvillei, Pteronotus quadridens, and Pteronotus portoricensis ) on Puerto Rico. Master's Theses and Doctoral Dissertations. Paper 349.
Schnitzler, H.-U., E. Kalko, I. Kaipf, and J. Mogdans. 1991. Comparative studies of echolocation and hunting behavior in the four species of Mormoopid bats of Jamaica. Bat Research News 32: 22-23.
Silva Taboada, G. 1979. Los murciélogos de Cuba. Habana. Editorial de la Academia de Ciencias de Cuba. pp. 425.
