


Taxonomic Status
Pteronotus parnellii was recognized in the 1980s to contain 9 subspecies, with the subspecies P. p. parnellii restricted to Jamaica and Cuba. However, in 2013
Clare et al. concluded from molecular, morphological, and acoustic data that parnellii is a cryptic species,
with four distinct species on the mainland which are different from the species occurring in the Greater Antilles. Pending final analyses, the species known as Pteronotus parnellii
will either be endemic to Jamaica or restricted to Jamaica and Cuba. As the type locality, Jamaica retains the name assigned by Gray in 1843.
Additional:
In Cuba, diet (from analyses of stomach contents) is dominated by moths (Lepidoptera). Orthoptera, Diptera, Coleoptera and Hemiptera are consumed less frequently (Silva Taboada 1979).
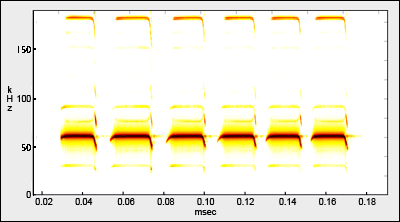
Considerable research has been conducted on the echolocation system and communication calls of Pteronotus parnellii. It is the only Mormoopidae in Jamaica and, indeed, the only bat in the New World, to use a constant-frequency (CF), as opposed to a frequency-modulated (FM), echolocation signal. CF calls allow this species to hunt insects (esp. moths) within a densely-cluttered forest (Schnitzler et al. 1991). CF echoes that are a consequence of Doppler effects associated with the fluttering wingbeats of insects enable this species to distinguish echoes of potential prey from echoes produced by cluttered vegetation (Kṏssl et al. 1999, 2003).
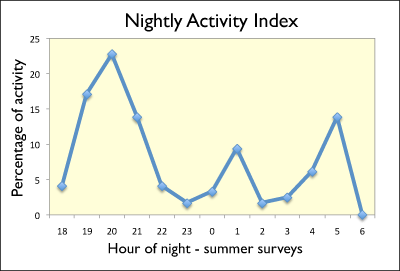
After departing from Windsor Great Cave shortly before sunset, Pteronouts parnellii commute along flyways associated with the vegetation patterns of the landscape: streams of P. parnellii can be seen crossing roads only where tree-crowns connect over the road. The most distant segment of the flyway we've identified thus far is located a linear 3.2 km north of the top opening of Windsor Great Cave; however, based on acoustic surveys we've conducted on the north coast, in the Coral Spring-Mountain Spring Protected Area (16 km northeast of Windsor), we suspect the flyway used by P. parnellii may be a considerable corridor for them.
Acknowledgements:
Data presented on the Windsor Great Cave colony were collected through the support of:
| |
|
|
Literature Reviewed
Clare, E.L., A.M. Adams, A. Z. Maya-Simoes, J.L. Eger, P. D.N. Hebert, and M.B. Fenton. 2013. Diversification and reproductive isolation: cryptic species in the only New World high-duty cycle bat,
Pteronotus parnellii. BMC Evolutionary Biology 2013, 13:26.
Emrich, M.A., E.L. Clare, W.O.C. Symondson, S.E. Koenig, and M.B. Fenton. 2013.
Resource partitioning by insectivorous bats in Jamaica. Molecular Ecology 23: 3648-3656.
Genoways, H.H., R.J. Baker, J.W. Bickham, C. J. Phillips. 2005. Bats of Jamaica. Museum Texas Tech Univ. Texas, USA. pp 155.
Goodwin, R.E. 1970. The ecology of Jamaican bats. J. Mammology 51: 571-579.
Kanwal, J.S., S. Matsumura, K. Ohlemiller, and N. Suga. 1994. Analysis of acoustic elements and syntax in communcation sounds emitted by mustached bats. Journal of the Acoustical Society of America 96:1229-1254.
Kṏssl, M., F Mayer, G. Frank, M. Faulstich, and I.J. Russell. 1999. Evoluationary adaptations of coclear function in Jamaican mormoopid bats. Journal of Comparative Physiology A: Sensory, Neural, and Behavioral Physiology 185:217-228.
Kṏssl, M., E. Foeller, M. Drexl, M. Vater, E. Mora, F. Coro, and I.J. Russell. 2003. Postnatal development of cochlear function in the Mustached Bat Pteronotus parnellii. Journal of Neurophysiology 90:2261-2273.
Mancina, C.A., L. Garcia-Rivera, and B.W. Miller. 2012. Wing morphology, echolocation, and resource partitioning in syntopic Cuban mormoopid bats. Journal of Mammology 95: 1308-1317.
Schnitzler, H.-U., E. Kalko, I. Kaipf, and J. Mogdans. 1991. Comparative studies of echolocation and hunting behavior in the four species of Mormoopid bats of Jamaica. Bat Research News 32: 22-23.
Silva-Taboada, G. 1979. Los murciélogos de Cuba. Habana. Editorial de la Academia de Ciencias de Cuba. pp. 425.