

Additional:
In our monitoring of the bat colony in Windsor Cave, we occasionally observe small, blood-filled ticks (Ixodida: Acarina) attached to individual Mormoops. This gives some insight into their flight or hunting behaviour, as clusters of ticks are most frequently encountered in the vegetation along the edges of trails or in pastures, less than two meters above the ground.
In Cuba, the diet of Mormoops blainvillei is dominated by Lepidoptera; true bugs (Hemiptera) and beetles (Coleoptera) also are occasionally consumed (Silva Taboada 1979). In Puerto Rico, Lepidoptera also represent the dominant component of the species' diet; Coleoptera and Hymenoptera (bees, wasps, ants) also are consumed occasionally; Hemiptera, Diptera (flies), and Araneae (spiders) rarely (Rolfe 2011).
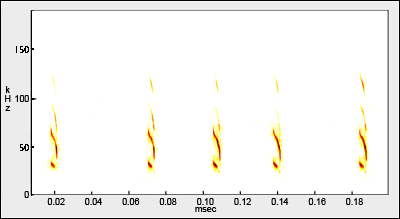
Although delicate in appearance, Mormoops has a remarkably swift flight and the vibration of the wings makes a distinctive humming “woosh” as they fly past (see also Goodwin 1970). You don’t need an acoustic bat detector to confirm Mormoops!
On Cuba, pregnant Mormoops blainvillei are found from March to June. In May the females begin roosting in different caves from the males, birthing begins mid-June, and lactating females are found into September (Silva-Taboada 1979).
In Windsor Cave, we've never captured any visibly- or palpably pregnant or lactating females, nor have we ever captured any juveniles. In the months when we commonly captured females (late July thru February), the female-to-male ratio was 1 : 2.6. We've never captured females in March. In April the capture ratio has been 1 (F) : 10 (M). Alas, we’re missing data for May and June.
More research is needed to understand the spatio-temporal movements of females in order to identify nursery caves in the wider landscape.
RESEARCHER ALERT: This species is extremely sensitive to being kept in cloth holding bags: the skin at the carpal bend can abrade completely and very quickly. Caution is needed when working with them.
Acknowledgements:
Data presented on the Windsor Great Cave colony were collected through the support of:
| |
|
|
Literature Reviewed
Emrich, M.A., E.L. Clare, W.O.C. Symondson, S.E. Koenig, and M.B. Fenton. 2013. Resource partitioning by insectivorous bats in
Jamaica. Molecular Ecology 23: 3648-3656.
Mancina, C.A., L. Garcia-Rivera, and B.W. Miller. 2012. Wing morphology, echolocation, and resource partitioning in syntopic Cuban mormoopid bats. J. Mammology 95: 1308-1317.
Goodwin, R.E. 1970. The ecology of Jamaican bats. J. Mammology 51: 571-579.
Rolfe, A.K. 2011. Diet of Three Mormoopid Bats (Mormoops blainvillei, Pteronotus quadridens, and Pteronotus portoricensis ) on Puerto Rico. Master's Theses and Doctoral Dissertations. Paper 349.
Schnitzler, H.-U., E. Kalko, I. Kaipf, and J. Mogdans. 1991. Comparative studies of echolocation and hunting behavior in the four species of Mormoopid bats of Jamaica. Bat Research News 32: 22-23.
Silva Taboada, G. 1979. Los murciélogos de Cuba. Habana. Editorial de la Academia de Ciencias de Cuba.